![[灵蜂智能官网]-专业红外热像仪厂家](http://linfun.com.cn/wp-content/uploads/2022/06/2022060807191642.png)
![[灵蜂智能官网]-专业红外热像仪厂家](http://linfun.com.cn/wp-content/uploads/2021/10/202110130602107.png)
研究者关注的巨嘴鸟是Ramphastos toco Muller,它是所有巨嘴鸟中具有最大喙子的。然而,至关重要的是,调节血流可以控制喙子的热交换。研究者检查了巨嘴鸟的喙子是否可以作为热量散失的热窗口,是否能够在热中性区域内和上方打开并闭合,以在较低温度下保存新陈代谢的热量。通过使用红外热像仪来研究环境温度(Ta)的变化对鸟类身体不同部位的热交换曲线的影响。
研究发现,巨嘴鸟的喙子通过陡峭的个体发育而达到其非凡的大小,导致喙子的表面积比根据关系预测的表面积大25至40倍。此后,当少年达到其成年体重的近80%时,该喙子就显示出更快的异速增长。
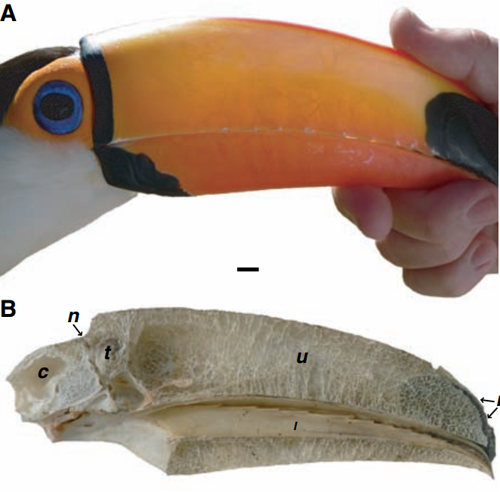
图为成人toco巨嘴鸟喙子和博物馆标本
通过红外热像仪技术观测后,正如预期的那样,羽毛覆盖区域的温度仅略高于环境温度,并且仅在最低温度下才发生显着变化;眼睛区域的温度几乎保持恒定。相比之下,在所有红外热像仪技术下,巨嘴鸟的体温均为38°至39°C。这表明不考虑红外热像仪技术,连续血液流向眼睛周围的裸露皮肤。近端和远端比羽毛区域变化更大,但主要趋势是在近端区域(成年鸟)中在20°至25°C之间出现较大的梯度,直到Ta高于25°时,远端区域才表现出持续的扩张。
因此,喙子的近端区域主要用于在较低的红外热像温度(>16°至25°C)下散发热量,并且随着温度的升高,远端区域开始接收增加的血流量,开始变暖,从而帮助禽类应对额外的热负荷。根据热敏窗的预期,喙子表面温度的最大变化发生在巨嘴鸟的热中性区(约18°至30°C)内。这些结果表明,在巨嘴鸟中,对喙子表面进行的血管舒缩调节具有温度调节功能。此外,在低于约21°C的空气温度下,呼吸频率和呼出的空气温度突然改变,会与喙子血管舒张的阈值平行。

图为巨嘴鸟在红外热像仪下两个不同的Ta(15°,20°)的表面温差的温度曲线。
通过红外热像仪技术观测后,当代谢热产生增加时,喙子的非凡的热交换能力也可能会发挥作用,例如在飞行过程中会发生热量产生是静止时的10到12倍时。对于一只鸟,喙子温度从约30°C到31°C开始,在飞行4分钟内开始升高,直到最终在第10分钟达到最高37°C。当热量产生增加时,喙子产生的热量损失可能对于维持足够的热量平衡至关重要。
因此,即使是2个月大的巨嘴鸟,喙子上的热量损失也很高,即使在寒冷的天气下也不能下调热量。巨嘴鸟在出生头3周内保持盲目和裸露状态,在4周时开始羽毛生长,仅在6周龄后才开始羽绒。即使在亚热带气候中,年轻的巨嘴鸟也将产生温度调节成本,由于它们控制喙子热交换的能力差而变得更具挑战性。
的表面温差的温度曲线1.png)
图为巨嘴鸟在两个不同的Ta(25°和35°C)的表面温差的温度曲线。
通过红外热像仪技术观察后成年喙子表面温度的时间变化迅速且可逆,在几分钟之内就发生了,这在研究者观察鸟类睡觉时最为明显。随着鸟类开始入睡,短暂的喙子血管舒张发生。由于鸟类与大多数吸热一样,会在夜间睡眠过程中降低体温,因此研究结果表明,随着热设定点的降低,巨嘴鸟有能力利用其喙子迅速排出体内热量。此外,还发现,熟睡的禽鸟显示出喙子表面温度的瞬时变化而没有觉醒的迹象,这表明与体温调节状态变化相关的睡眠状态转变。
最终结果表明,在理解巨嘴鸟的分布,生态和行为时,应考虑到热交换的限制以及喙子作为体温调节器官的潜在用途。考虑到喙子结构的快速辐射和鸟类喙形的多样性,喙子热损失造成的热量限制可能被证明是许多禽类动物的共同特征。
参考资料:
Glenn J. Tattersall, Denis V. Andrade, Augusto S. Abe. Heat Exchange from the Toucan Bill Reveals a Controllable Vascular Thermal Radiator. Science. 325:468-470, 2009.